
Developmental Biology
- Axon Guidance and Neuron Migration
- Cardiac Stem Cell Markers
- Cardiovascular System Development
- Digestive System Development
- Excretory System Development
- Gametes and Fertilization
- Gastrulation and Germ Layer Formation
- Immune System Development
- Morphogens, Embryonic Patterning and Axis Formation
- Muscular System Development
- Nervous System Development
- Neural Stem Cells
- Primordial and Mature Germ Cells
- Skeletal System Development
- Tissue Regeneration
- Wnt Family
The development of a new life is a spectacular process and represents a masterpiece of temporal and spatial control of gene expression. Developmental genetics studies the effect that genes have in a phenotype, given normal or abnormal epigenetic parameters. The findings of developmental biology can help to understand developmental abnormalities such as chromosomal aberrations that cause Down syndrome. An understanding of the specialization of cells during embryogenesis has provided information on how stem cells specialize into specific tissues and organs. This information has led, for example, to the cloning of specific organs for medical purposes. Another biologically important process that occurs during development is apoptosis—programmed cell death or "suicide." Many developmental models are used to elucidate the physiology and molecular basis of this cellular process.The development of life is a surprising process and is regulated under control of a series of temporal and spatial gene expressions. Developmental biology focuses on the study of the process. In animals, developmental events can be found in embryonic life, regeneration, asexual reproduction, metamorphosis and so on. The main processes of embryonic development include cell differentiation, growth, morphogenesis, regional specification and the overall control of timing. In plants, developmental events are found in embryos, vegetative reproduction and growth of roots, shoots and flowers. Thus, developmental biology study is associated with other fields such as genetics, cell biology and molecular biology and they promote each other.
Results of normal developmental biology studies can help to solve and understand some problems. For example, knowledge of apoptosis process helps to design cancer therapeutic strategies. The study of animal developmental biology can help create kinds of animal models to research human diseases and explain the physiology and molecular basis. The studies about risks from substances also help protect the fetus from damages. The cloning of organs for medical purposes is a result of specialization of cells during embryogenesis. Creative-Biomart is able to provide amounts of developmental biology related molecular tools for developmental biology studies.
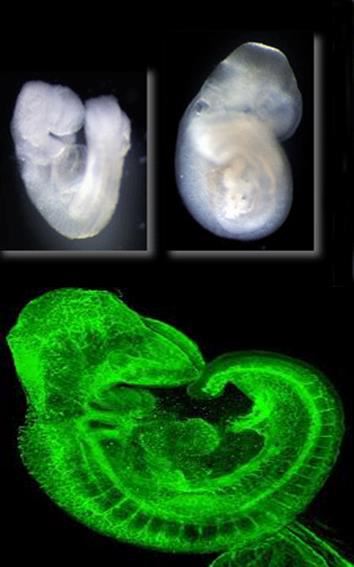 Figure 1. Developing life.
Figure 1. Developing life.Introduction
Developmental biology is one of the important basic branches of biological sciences. The content of research and the interdisciplinary content of many disciplines are interrelated and interrelated, especially with genetics, cell biology and molecular biology. It uses modern science and technology to study the processes and mechanisms of the analysis of organisms at the molecular, submicroscopic and cellular levels.
The discipline of studying the mechanisms of ontogeny by molecular biology and cell biology. It was developed from experimental embryology. Experimental embryology is to study the interrelationships between the various parts of the developing embryo and how they interact with each other. Developmental biology is to investigate what is the substance of this relationship, what substance (or substances) are at work, How the functioning substance differentiates the embryonic cells in a certain direction, how the differentiated cells constitute tissues or organs, to ensure the development of tissues and organs, how the normally developing embryos grow, mature, and become a growing individual, the latter developing Why do you gradually move toward aging after a certain period of time, and how to complete the individual development in the specified time and space sequence.
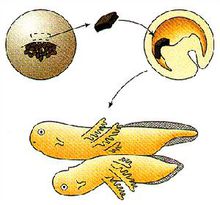 Figure 2. Process of cell differentiation.
Figure 2. Process of cell differentiation.Influences
As one of the most active fields of contemporary life science research, developmental biology brings together various disciplines such as molecular biology, cell biology, genetics, immunology, embryology, evolutionary biology and ecology. Use to reveal the essential laws of life development; On the other hand, developmental research has existed in various fields of biology and has become an essential element of other disciplines. The development of developmental biology research will certainly promote the development of other disciplines. Therefore, developmental biology is one of the most important basic disciplines. Developmental biology is closely related to the use of medical and health, agricultural production and the use of biological resources, such as the mechanism of fertilization and early embryo development, the mechanisms of tumors, AIDS, malformation, and the mechanism of aging, for family planning, prenatal and postnatal care, and healthy living. And agriculture, forestry, animal husbandry and other production have a profound impact.
Conclusions
The results of normal developmental biology studies can help solve and understand certain problems. For example, understanding the process of apoptosis helps to design cancer treatment strategies. Research in animal developmental biology can help create a variety of animal models to study human diseases and explain physiological and molecular basis. Research on substance risks also helps protect the fetus from damage. Organ cloning for medical purposes is the result of cell specialization during embryogenesis. Creative-Biomart provides a wealth of molecular tools related to developmental biology for developmental biology research.
Reference:
- Reddien P.W.; et al. Fundamentals of planarian regeneration. Annual Review of Cell and Developmental Biology. 2004, 20: 725–757