Uncategorized Thursday, 2016/06/16
Newest article from Science Magazine: http://science.sciencemag.org/content/352/6292/1390 A single zygote with a defined DNA sequence gives rise to all the varied cells of the mammalian body. This variety of cell fates may arise in part from cell-to-cell variability in the gene expression programs of embryonic stem cells (ESCs). Three recent studies have taken different approaches to characterizing this variability in gene expression in stem cells. These results suggest that microRNAs (miRNAs) could play an important role in controlling and generating this variability. Pluripotent stem cells move irreversibly down specific differentiation pathways during development. Yet the effective concentrations of key pluripotency factors fluctuate over time, and seemingly uniform populations of stem cells can exhibit multiple cell fates. Differentiated cells can also be driven to dedifferentiate, as seen in induced pluripotent stem cells (iPSCs). Thus, cells may exist in a continuum of “cell states” that can interconvert with each other and have varying potential to differentiate or to self-renew. However, the factors that drive interconversion of cell states have remained poorly defined, and variability is often attributed to “noise”—the random fluctuations of biochemical events. This raises the question of whether such variability is stochastic or coherently organized.
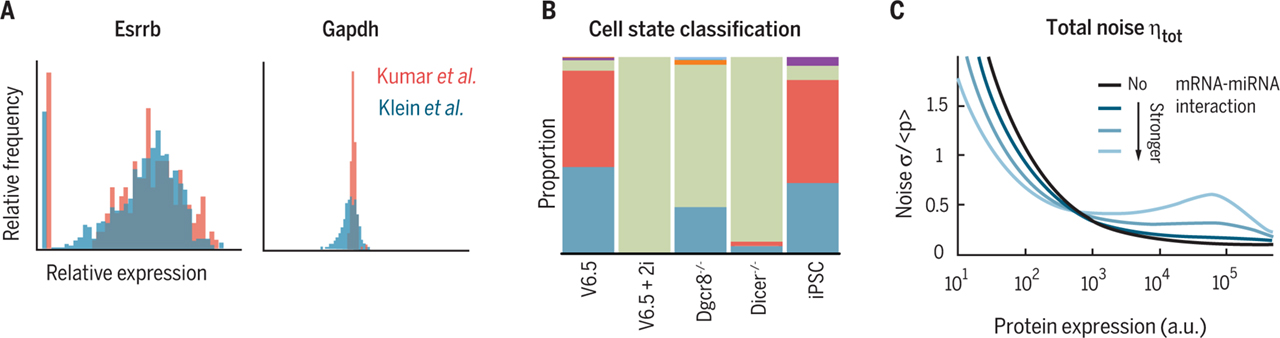 Fig.1 (A) Comparing the single-cell RNA sequencing data of Kumar et al. and Klein et al. reveals that the pluripotency factor Esrrb is heterogeneously expressed, whereas Gapdh is narrowly expressed. (B) Embryonic stem cells under chemical (2i) and genetic (Dgcr8−/−, Dicer−/−) perturbations show similar cell states (green, ground; blue, transition 1; red, transition 2; purple, primed; orange, fibroblast states) within the total population relative to the wild type (V6.5). (C) In the model proposed by Schmiedel et al., at low protein expression (a.u., arbitrary units), miRNA activity decreases gene expression variability (noise = standard deviation/mean); at high protein expression, miRNA activity increases gene expression variability through transmission of variability in the miRNA pool.
Fig.1 (A) Comparing the single-cell RNA sequencing data of Kumar et al. and Klein et al. reveals that the pluripotency factor Esrrb is heterogeneously expressed, whereas Gapdh is narrowly expressed. (B) Embryonic stem cells under chemical (2i) and genetic (Dgcr8−/−, Dicer−/−) perturbations show similar cell states (green, ground; blue, transition 1; red, transition 2; purple, primed; orange, fibroblast states) within the total population relative to the wild type (V6.5). (C) In the model proposed by Schmiedel et al., at low protein expression (a.u., arbitrary units), miRNA activity decreases gene expression variability (noise = standard deviation/mean); at high protein expression, miRNA activity increases gene expression variability through transmission of variability in the miRNA pool.
Kumar et al. used single-molecule RNA fluorescence in situ hybridization (FISH), single-cell reverse transcription polymerase chain reaction (RT-PCR), immunofluorescence, and single-cell RNA sequencing to explore the gene variability landscape of pluripotent ESCs. Klein et al. performed deep RNA sequencing using a dropletmicrofluidic approach to identify genes displaying variable transcript levels among single ESCs. Both reports find that the majority of genes, such as those encoding housekeeping and metabolic factors, display stable normal distributions of expression. However, RNA levels for a subset of genes, such as signaling factors and developmental regulators, are more variable cell-to-cell and display bimodal or heterogeneous expression patterns. Among these variable genes are pluripotency factors known to fluctuate cell-to-cell across the population (e.g., Nanog, Esrrb) and whose expression is key to pluripotency. Strikingly, for many genes, both Kumar et al. and Klein et al. report similar distribution of expression and extent of variability (see the figure, panel A). Determining the set of genes with variable expression within an ESC population represents an important step in understanding what drives the interconversion between cell states. A cell state can be defined as the relatively stable expression of a group of genes at (or near) specific levels. Both Kumar et al. and Klein et al. undertook principal components analysis to identify gene expression relationships. In the case of Kumar et al., the range of cell types analyzed included ESCs lacking mature miRNAs and lacking signaling through the ERK/GSK3 kinase pathway (see the figure, panel B). Similar discrete cell states were observed across the various experimental conditions, although the portion of cells in each cell state varied depending on the nature of the perturbation. For example, inhibiting the ERK/GSK3 pathway (the 2i condition) or removing miRNA regulation (Dicer−/−, Dgcr8−/−) resulted in a common “ground” state with little variation in the expression of key pluripotency genes (green in the figure, panel B). Other ESCs were classified into “transition” states according to their expression of key components (red and blue in the figure, panel B). Labeling these as transition states is reasonable given the overlap in expression with the ground state, but the experimental methodologies did not allow testing of the interconversion of individual cells between states. Klein et al. identified a similar continuum of heterogeneous cell states within wild-type ESC populations. Further, they examined gene expression covariation to identify a set of genes closely correlating with a given gene and with each other to infer regulatory information. This topological network analysis of covariation patterns reveals that pluripotency factors such as Nanog are strongly enriched for interaction with other pluripotency factors, which in turn form a gene neighborhood. Thus, pluripotency factors likely fluctuate together as a network to generate transitions between cell states. The third study, by Schmiedel et al., may provide a key insight. Combining a reporter construct for monitoring miRNA activity with theoretical and mathematical modeling, they proposed that miRNAs increase protein expression noise for highly expressed genes, and that they decrease such noise for genes that are not highly expressed (see the figure, panel C). Reduction in gene expression noise was previously proposed as a hallmark of miRNA function in the context of transcriptional dynamics. Yet viewed in the context of findings from single-cell RNA sequencing in ESCs, a role for miRNAs in generating gene expression variability is noteworthy. Perhaps miRNA activity increases the cell-to-cell variability of key pluripotency factors through the transmission of variability in the miRNA pool itself. This is consistent with the cell-to-cell variation in miRNA activity observed by Schmiedel et al. . Variability is then transmitted throughout the pluripotency network according to the topology of gene regulatory interactions. Downstream action by pluripotency factors may then establish miRNA expression patterns through a virtuous cycle that determines a cell state. Because miRNAs have the potential to control expression of large numbers of genes, variability in miRNA activity would provide a potential mechanism for generating transitions between cell states. The three studies raise more questions than they answer. For example, it is not known what accounts for the network of genes that covary between cell states, nor the process that produces variability in key pluripotency genes. Variation in the expression of miRNAs may provide one answer but raises the issue of what accounts for miRNA variation. Further, it is not clear whether similar mechanisms are important for the activity of other stem cells, such as cancer stem cells. A major limitation of all three studies is the inability to follow variation within individual cells and relate this to their biochemical state. Nonetheless, if correct, miRNA-guided variability will have implications for intrinsically generated heterogeneity in tumorigenesis, development, and immune function. [table caption="Related Products" width="800" colwidth="80|180|180" colalign="left|left|center|center|center"] No.,Product classify,Representative Product 1,Lineage Markers Proteins,Recombinant Mouse Notch1 protein 2,Embryonic Stem Cells Escs Proteins,Recombinant Mouse STAT3 Protein 3,Cancer Stem Cells Cscs Proteins,Recombinant Mouse CXCR4 Protein 4,Stem Cell Signaling Pathways Proteins,Recombinant Human WNT5A protein [/table]