
Wingless-INT (WNT)

🧪 WNT3-5160M
Source:
Species: Mouse
Tag: Non
Conjugation:
Protein Length:
🧪 WNT11-3737H
Source: E.coli
Species: Human
Tag: GST
Conjugation:
Protein Length: 1-354aa
🧪 WNT3-3738H
Source: E.coli
Species: Human
Tag: His
Conjugation:
Protein Length: 1-355aa
🧪 WNT4-3740H
Source: E.coli
Species: Human
Tag: His
Conjugation:
Protein Length: 1-351 aa

🧪 WNT8B-3741H
Source: E.coli
Species: Human
Tag: GST
Conjugation:
Protein Length: 197-267aa
🧪 Wnt5a-2055M
Source: E.coli
Species: Mouse
Tag: GST&His
Conjugation:
Protein Length: Ile62~Lys380
🧪 WNT4-517HFL
Source: HEK293
Species: Human
Tag: Fc&Flag
Conjugation:
Protein Length: Full Length
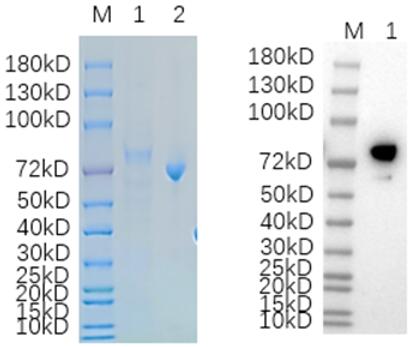
🧪 WNT3A-3739H
Source: E.coli
Species: Human
Tag: His
Conjugation:
Protein Length: 19-120aa








SubCategories
Wnt Superfamily—Product Overview
The Wnt protein family consists of highly conserved secreted signaling molecules that play fundamental roles in embryonic development, cell proliferation, and tissue homeostasis. Through activation of canonical and non-canonical Wnt signaling pathways, these proteins regulate processes such as cell fate determination, migration, and polarity. Dysregulation of Wnt signaling is implicated in numerous diseases, including cancer and degenerative disorders. Creative BioMart offers a comprehensive range of recombinant Wnt family proteins and related products to support research in developmental biology, oncology, and regenerative medicine.
Jump to Section
Product Families
-
Wnt Proteins
-
Regulatory Protein
-
Receptors and Associated Proteins
Background
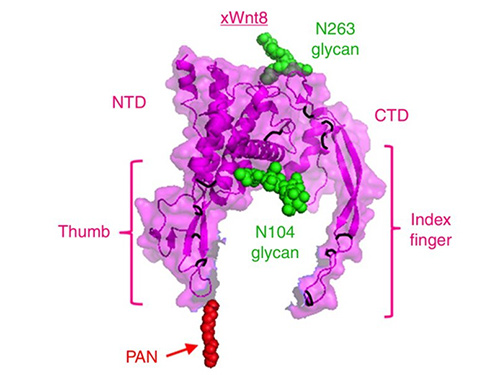
Wnt proteins are secreted glycoproteins (~350-400 amino acids) with ~22 conserved cysteines forming multiple intramolecular disulfides. They contain N-terminal signal peptides and undergo extensive ER processing (N-glycosylation and lipid acylation by Porcupine). These modifications render Wnts hydrophobic and membrane-associated. For example, mammalian Wnt1 has several N-linked glycans, whereas Drosophila WntD lacks them. The crystal structure of Xenopus Wnt8 revealed a unique two-domain fold (helical N-terminal domain with 10 cysteines and β-sheet C-terminal domain with 6 cysteines). Wnts engage target cells by binding to the cysteine-rich extracellular domain of Frizzled receptors.
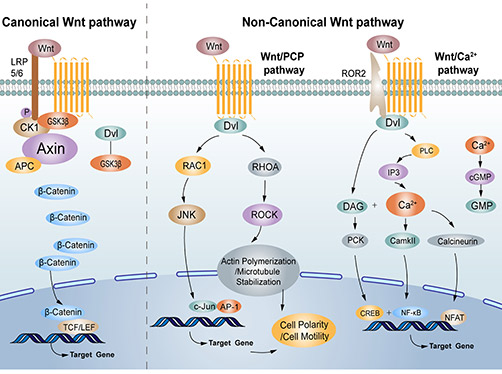
Wnt signaling is divided into canonical (β-catenin-dependent) and non-canonical branches. In the canonical pathway, Wnt ligands bind Frizzled and LRP5/6 co-receptors, leading to Dishevelled-mediated inhibition of the Axin/APC/GSK-3β "destruction complex". Stabilized β-catenin accumulates and translocates to the nucleus, activating TCF/LEF-driven target genes. Non-canonical Wnts (e.g. Wnt5a, Wnt11) signal through β-catenin-independent pathways (planar cell polarity or Ca2+-mediated pathways) that regulate cytoskeletal polarity and cell migration.
Learn More
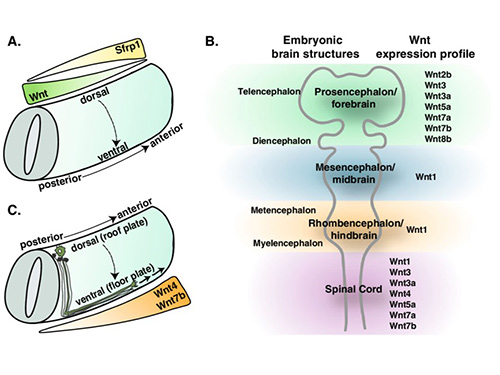
Wnts are key embryonic morphogens. They specify body axes and tissue fates across species: high Wnt/β-catenin specifies posterior (mesendoderm) and low Wnt allows anterior (neural) differentiation. For example, murine Wnt3a is essential for gastrulation: Wnt3a-null embryos fail to form posterior mesoderm and instead become neural tissue. Similarly, Drosophila Wingless patterns segment polarity (wingless mutants lack defined segments). Overall, Wnt gradients orchestrate the development of diverse structures (CNS, gut, limbs, etc.) by coordinating proliferation, differentiation and polarity.
LEARN MORE
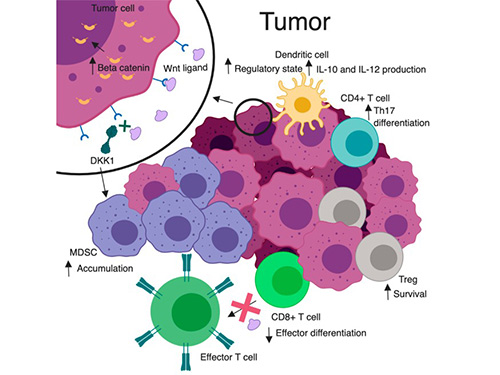
Aberrant Wnt signaling is a hallmark of many diseases, particularly cancer. Constitutive Wnt/β-catenin activation drives proliferation and stemness. Truncating APC (stabilizing β-catenin) mutations occur in ∼85% of colorectal tumors, and other tumors often harbor activating β-catenin or Axin mutations. Dysregulated Wnt/β-catenin promotes tumorigenesis in colorectal, liver, melanoma and other cancers. Wnt pathway defects also underlie fibrotic, metabolic and skeletal disorders. Thus, dysregulation of the Wnt pathway is a central driver of multiple pathologies.
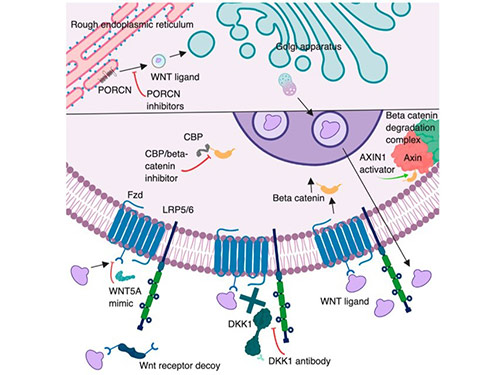
Components of the Wnt pathway are active drug targets in oncology and regenerative medicine. Strategies include inhibitors of Wnt secretion (e.g., porcupine acyltransferase blockers) and ligand traps/antibodies that sequester Wnts or block Frizzled. For example, LGK974 (a PORCN inhibitor) potently blocks Wnt ligand maturation and shows tumor suppressive activity in preclinical models. Conversely, enhancing Wnt activity has clinical applications: anti-sclerostin antibodies are FDA-approved to increase bone formation via Wnt signaling. In summary, numerous Wnt-targeting small molecules and biologics are in development or clinical trials for cancer, bone and regenerative applications.
Applications

Regenerative Medicine
Wnt proteins play a key role in regulating stem cell proliferation, differentiation and tissue regeneration. Recombinant Wnts are used to promote repair of bone, skin and neural tissues. Their controlled application in regenerative therapies offers significant potential for accelerating healing and restoring function to damaged organs.
Cancer Research
Aberrant activation of Wnt signaling has been implicated in the pathogenesis of several cancers, including colorectal, breast and melanoma. Both native and recombinant Wnt proteins serve as critical tools for elucidating tumor biology. Targeting Wnt pathways is a promising strategy for developing anti-cancer therapeutics and understanding mechanisms of tumor progression.
Drug Discovery and Development
Drug Discovery and Development Recombinant Wnt proteins are widely used in high-throughput drug screening platforms to identify modulators of Wnt signaling. These applications aid in the discovery of therapeutic agents that can either enhance tissue regeneration or inhibit pathological Wnt activity, particularly in oncology and fibrotic diseases.
Organoid and 3D Culture Systems
The incorporation of Wnt proteins is essential for the successful generation and maintenance of organoids, such as intestinal, liver and neuronal models. Recombinant Wnts provide critical niche signals that allow organ-specific stem cells to self-organize, providing powerful systems for disease modeling, drug testing and personalized medicine research.
Cosmetic and Anti-Aging Applications
Modulation of Wnt signaling has been investigated for applications in dermatology and aesthetic medicine, including hair follicle regeneration and skin rejuvenation. Recombinant Wnt proteins and Wnt pathway activators are being explored to develop novel treatments to promote tissue renewal and combat age-related degeneration.
Veterinary and Agricultural Biotechnology
In veterinary medicine, recombinant Wnt proteins are used to enhance tissue repair and regeneration processes in farm animals. In addition, studies of Wnt-like signaling mechanisms in plants aim to improve growth and developmental patterns, offering promising biotechnological strategies for increasing agricultural productivity.
Product Features
-
Broad Selection: Comprehensive catalog covering key Wnt isoforms (e.g., Wnt1, Wnt3a, Wnt5a) in both human and murine formats, available in high-purity recombinant and native forms to support a wide range of research applications.
-
High Purity and Activity: All Wnt proteins are rigorously tested for bioactivity using validated cell-based assays, ensuring consistent performance in signaling studies, stem cell maintenance, and regenerative models.
-
Superior Stability and Bioavailability: Optimized formulations with proprietary stabilization techniques to maintain protein conformation, solubility, and signaling potency, minimizing degradation during storage and experimental use.
-
Custom Services: Flexible production options to meet both small-scale research and large-scale commercial needs, with custom concentrations, buffer compositions, and bulk packaging available upon request.
-
Ready-to-Use: Pre-formulated Wnt solutions and media supplements tailored for rapid integration into organoid culture workflows, ensuring reproducibility and efficiency in 3D tissue modeling.
-
Expert Technical Support: Dedicated scientific team providing protocol optimization advice, troubleshooting assistance, and custom development services to help researchers maximize the value of their Wnt-based experiments.
Case Study
Case 1: Wnt8 Localization Mechanisms
Suzuki et al., 2024.Dissection of N-deacetylase and N-sulfotransferase activities of NDST1 and their effects on Wnt8 distribution and signaling in Xenopus embryos.
Wnt proteins regulate key cellular processes, including stem cell maintenance, cancer development, and embryonic patterning. Wnt8 distribution on the cell surface depends on specific modifications of heparan sulfate (HS) proteoglycans. In Xenopus embryos, N-sulfation (but not deacetylation) by NDST1 is crucial for Wnt8 localization and signaling, as revealed by a novel NDST1 mutant that selectively affects HS chain modifications.
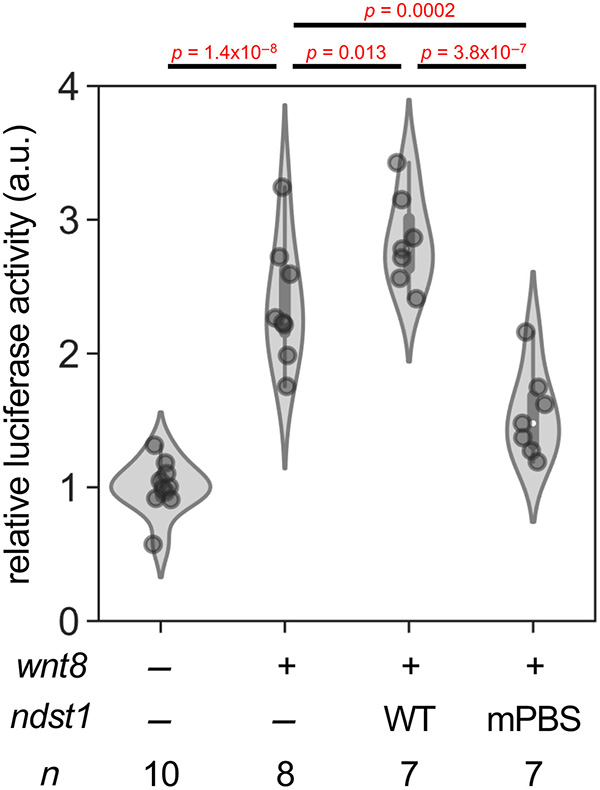
Figure 1. Canonical Wnt signaling required N-sulfation-rich HS chains rather than N-deacetylation-rich HS chains. TOP-Flash reporter assay. Amounts of mRNAs: wnt8, 100 pg/embryo; ndst1 (WT or mPBS), 500 pg/embryo. Amount of TOP-Flash reporter DNA: 100 pg/embryo.
Case 2: Wnt3 in 5FU-Resistant OSCC
Zhang et al., 2024.Wnt3 promotes chemoresistance to 5-Fluorouracil in oral squamous cell carcinoma via activating the canonical β-catenin pathway.
5-Fluorouracil (5FU) resistance limits treatment of oral squamous cell carcinoma (OSCC), prompting investigation into the underlying mechanisms. This study found that activation of the Wnt/β-catenin pathway, specifically through upregulation of Wnt3, promotes 5FU resistance. Knockdown of Wnt3 or treatment with the Wnt inhibitor MSAB restored 5FU sensitivity, suggesting that targeting Wnt3 may be a novel strategy to overcome chemoresistance in OSCC.
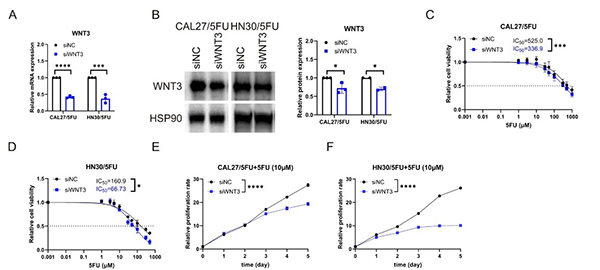
Figure 2. Downregulation of Wnt3 enhances 5FU efficacy in 5FU-resistant OSCC cells. A, B RT-qPCR and western blot evaluations of Wnt3 expression post-knockdown in 5FU-R cells. C, D Decreased IC50 of 5FU in 5FU-R cells after Wnt3 knockdown. E, F Inhibited proliferation under 5FU exposure in 5FU-R cells after transfection with siWnt3.
FAQs
-
Q: What types of Wnt proteins do you offer?
A: We provide a comprehensive portfolio of recombinant Wnt proteins, including key isoforms such as Wnt1, Wnt3a, Wnt5a, Wnt7a, and Wnt8, among others. Our products are available in human and murine forms and are produced in both mammalian and insect cell expression systems to ensure correct folding and bioactivity. -
Q: How do you validate the biological activity of your Wnt proteins?
A: All of our Wnt proteins undergo rigorous bioactivity testing using validated cell-based assays. We ensure that each batch meets strict functional and purity criteria so that you can rely on consistent, reproducible results.
-
Q: What are the recommended storage conditions for your Wnt proteins?
A: Our Wnt proteins are supplied in lyophilized or stabilized liquid formats. Lyophilized products should be stored at -20°C or -80°C upon receipt, while liquid formulations are best stored at -80°C for long term storage. Detailed storage and handling instructions are included with each product to ensure optimal stability.
-
Q: Do you offer bulk supply or custom formulations?
A: Yes, we specialize in flexibility. We offer bulk quantities for large-scale projects and can provide custom concentrations, buffer systems, or formulation adjustments to meet your specific research or manufacturing needs. Contact our technical support team to discuss your custom project requirements. -
Q: Are your Wnt proteins suitable for clinical or translational research?
A: Certain Wnt proteins are available in research grade and cGMP compliant formats. If you require cGMP-grade material for cell therapy development, regenerative medicine or translational applications, please specify in your inquiry and we will guide you through the available options.
-
Q: How should I reconstitute the lyophilized Wnt proteins?
A: We recommend reconstituting lyophilized Wnt proteins with sterile water or a specified buffer containing 0.1% BSA to prevent protein adsorption and maintain bioactivity. Specific reconstitution instructions tailored to each product are provided in the product datasheet. -
Q: Can I use your Wnt proteins for organoid culture?
A: Absolutely. Our Wnt proteins are widely used in organoid and 3D culture systems. Their high biological activity supports robust organoid formation, stem cell maintenance, and tissue modeling. We also offer ready-to-use formulations specifically optimized for organoid protocols.