Protein SUMOylation Assay
Creative BioMart provides protein SUMOylation assay system for generating SUMOylated proteins in vitro. We provide carboxy-terminal SUMO-1, SUMO-2 or SUMO-3 peptide substrates which are linked covalently to specific lysine residues on the target protein via isopeptide bonds.
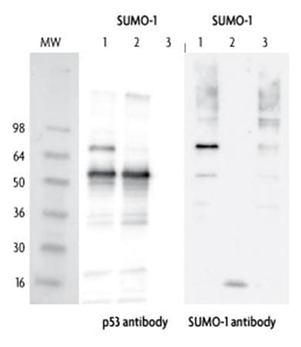
Western blot analysis of in vitro SUMOylation of p53 protein by wild-type and mutated isoforms of SUMO-1: The two Western blots were incubated with p53 antibody (1:5000 dilution) and SUMO-1 antibody (1:4000 dilution). The p53 protein is SUMOylated only by the wild-type SUMO-1 protein.
Applications
- Investigation of SUMOylation effects on transcription factor activity
- Understanding the role of SUMOylation in the regulation of cellular processes
- Identification of novel proteins as targets for SUMO
Please get in contact with Creative BioMart for more information or a detailed discussion.
Contact us or send an email at for project quotations and more detailed information.
Quick Links
-

Papers’ PMID to Obtain Coupon
Submit Now -

Refer Friends & New Lab Start-up Promotions