
Enzyme Activity Assay
Background
Enzymes are the molecular powerhouses behind nearly all biological reactions, driving essential processes with remarkable specificity and efficiency. They play a central role in metabolism, cellular signaling, regulation, molecular transport, and even movement. At their core, enzymes catalyze the transformation of substrates into chemically modified products—whether by cleaving proteins (proteolysis), breaking down sugars (glycolysis), or adding functional groups through phosphorylation, acylation, glycosylation, and redox reactions. When enzyme activity goes awry, it can trigger or exacerbate disease, making these versatile catalysts prime targets for therapeutic intervention and drug development.
At Creative BioMart, we leverage decades of expertise in designing and manufacturing active enzymes and their substrates to offer an extensive range of biochemical assays. Our services are tailored to support scientists engaged in enzyme and protein research, providing robust solutions for various experimental needs.
What We Offer?
Service Offering
Enzyme Activity and Inhibition Kinetics
We specialize in evaluating the kinetic parameters of enzyme activity and inhibition. This service helps researchers understand the rate at which enzymes catalyze reactions and how inhibitors affect these processes.
Inhibitor Screening and Validation
Our comprehensive inhibitor screening services are designed to identify potential inhibitors of enzymes. We also validate these inhibitors through rigorous testing to ensure their efficacy and specificity.
IC50 Determination of Drug Compounds
We provide accurate determination of IC50 values for drug compounds, which is crucial for assessing their inhibitory potency against target enzymes.
Mode of Action Studies
We conduct detailed studies to analyze the mode of action of enzyme-inhibitor interactions. This helps in elucidating the mechanisms underlying enzyme inhibition.
Reversibility of Enzyme Inhibition
Our investigations into the reversibility of enzyme inhibition provide insights into the stability and dynamics of enzyme-inhibitor complexes.
Substrate Profiling
We offer substrate profiling services to analyze the selectivity of enzymes. This helps in understanding the range of substrates that an enzyme can utilize, providing valuable information for functional studies.
Our Distinct Assay Approach—Immunoassay-Based Enzyme Activity Measurement
At the core of our enzyme assays lies a highly selective method that combines immunocapture with functional enzyme readout.
Step-by-Step Overview
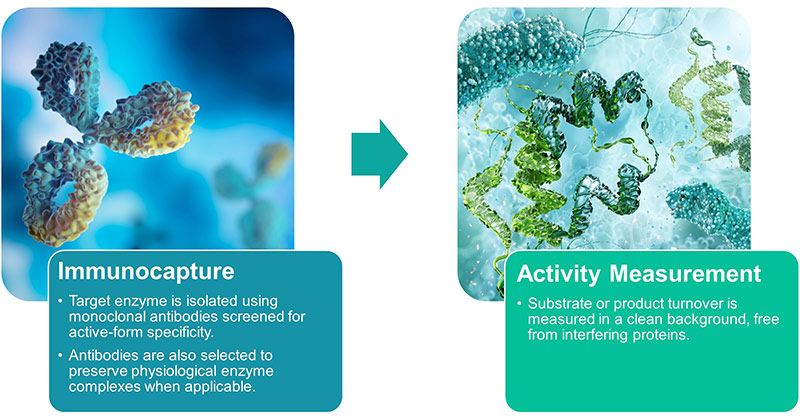
Key Advantages
- High Specificity: Eliminates interference from enzymes sharing substrates (e.g., ATP, NADH, pyruvate).
- Physiological Relevance: Maintains native enzyme complexes as seen in situ.
- Multiplex Compatibility: Supports downstream analysis such as mass spectrometry for post-translational modifications (PTMs).
- Dual-Mode Analysis: Combine activity assays with our Sandwich ELISA kits to calculate specific activity (activity per protein quantity).
Why Choose Us?
- 10+ Years of Assay Innovation: We bring decades of expertise in enzyme biochemistry, assay design, and substrate development to every project.
- Proprietary Monoclonal Antibody Platform: Our in-house antibody discovery pipeline delivers ultra-specific binders that isolate only the catalytically active form of the enzyme.
- Publication-Ready Data: High-quality data outputs are formatted and validated for seamless integration into publications, patents, or regulatory submissions.
- Custom Assay Development: Need something tailored? We build assays from the ground up, optimized to your target enzyme, matrix, and research goals.
- Broad Assay Portfolio: We support over 100 enzyme targets spanning kinases, phosphatases, proteases, glycosidases, and more.
- High-Throughput Compatibility: Scalable platforms support 96- and 384-well formats for screening campaigns and large-scale studies.
Case Study
* NOTE: We prioritize confidentiality to safeguard our clients’ technology and intellectual property. As an alternative, we present selected published research articles as representative case studies. For details on the assay services and products used in these studies, please refer to the relevant sections of the cited literature.
Case 1: Apigenin inhibits pepsin activity via direct binding and structural modulation
Guo et al., 2023. doi:10.1016/j.saa.2022.122281
This study investigated how apigenin (AP), a dietary polyphenol, interacts with pepsin and affects its enzymatic activity. Using spectroscopic techniques and molecular docking, researchers found that AP binds spontaneously to a single site on pepsin, primarily through hydrogen bonding and van der Waals forces. This binding caused conformational changes in the enzyme, confirmed by fluorescence analysis. Functionally, AP inhibited pepsin’s activity, while pepsin reduced AP’s antioxidant capacity. These findings highlight the mutual modulation between AP and pepsin at the enzymatic level.
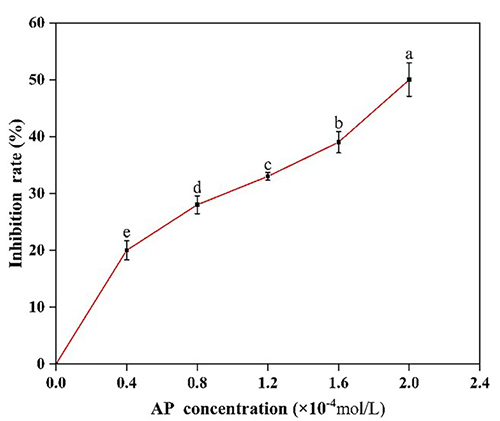
Figure 1. Effects of AP on the inhibition rate of pepsin; AP, short for apigenin. Different letters on the curve indicate differences among samples. (Guo et al., 2023)
Case 2: Role of Arg152 in GPX4 activity and its implications in SSMD
Roveri et al., 2023. doi:10.1016/j.redox.2023.102806
This study explored the biochemical impact of the Arg152His mutation in Glutathione Peroxidase 4 (GPX4), linked to Sedaghatian-type Spondylometaphyseal Dysplasia (SSMD). While the mutation did not alter the enzyme’s catalytic mechanism or general kinetics, differences emerged in the presence of cardiolipin-containing liposomes. Wild-type GPX4 showed positive cooperativity and enhanced activity due to surface-sensing behavior near the Arg152 site. This cardiolipin-dependent regulation was lost in the mutant, suggesting a disrupted physiological role of GPX4 in mitochondria relevant to SSMD pathology.
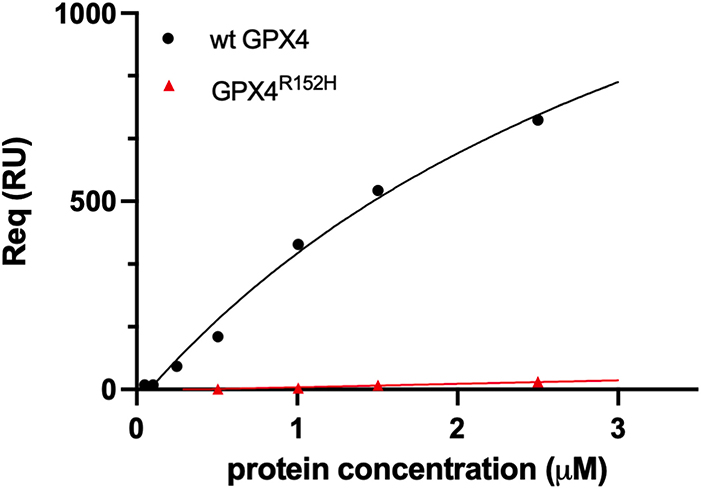
Figure 2. SPR analysis of wild-type or GPX4R152H binding over TOCL and DOPC bilayers. (Roveri et al., 2023)
Customer Testimonials
-
“We’ve used Creative BioMart’s enzyme activity assays extensively to evaluate enzyme kinetics and screen for inhibitors. The quantitative data—especially from initial velocity measurements and dose-response curves—helped us rapidly shortlist viable compounds. Their technical team was responsive during assay optimization, particularly in tailoring conditions for our serine protease targets.”
— Research Scientist | Biopharma Company
-
“Our enzyme-inhibitor interaction studies greatly benefited from Creative BioMart’s mechanistic assay services. Their reversible vs. irreversible binding analysis using pre-incubation and dilution methods provided critical insights into compound stability. The data quality supported confident decision-making for our lead candidate selection in early-stage drug discovery.”
— Senior Scientist | Biotechnology Company
-
“For a recent academic study on substrate selectivity, we turned to Creative BioMart’s enzyme activity assays. Their substrate profiling was thorough, using a diverse panel that revealed unexpected enzyme preferences. The immunoassay-based detection ensured high specificity, minimizing background from off-target proteins—a crucial factor for our signaling enzyme research.”
— Principal Investigator | Academic Medical Center
-
“In our drug discovery program, IC50 determination is essential, and Creative BioMart has become our go-to resource. Their accurate, reproducible data—validated using controls and multiple replicates—enabled efficient ranking of compound potency. The integration of ELISA-based quantitation with enzymatic activity data provided specific activity metrics that fed directly into our SAR analysis.”
— Lead Scientist | Pharmaceutical Research Group
-
“We’ve collaborated with Creative BioMart for over a year on enzyme assay development. Their immunocapture-based approach ensures we’re measuring the active form of our recombinant enzymes. Additionally, their capacity to assess post-translational modifications like phosphorylation status has added functional depth to our data. Their technical staff has consistently provided insightful input throughout the process.”
— Research Director | Industrial Biotech Organization
FAQs
-
Q: What makes your enzyme activity assays more reliable than others?
A: Our enzyme activity assays are designed with precision and reliability in mind. We use a unique immunoassay-based approach that involves capturing the enzyme of interest with highly specific monoclonal antibodies. This ensures that only the active form of the enzyme is measured, eliminating potential interference from other proteins that might use or produce the same substrates or products. Additionally, our assays are validated through rigorous testing, providing accurate and reproducible results. -
Q: How do you ensure the enzyme is in its active form during the assay?
A: We carefully screen our monoclonal antibodies to isolate the enzyme in its active form. This ensures that the enzyme is functional and representative of its natural state in situ. By capturing the enzyme in this way, we can provide reliable data on its activity, without interference from inactive or denatured forms of the enzyme. -
Q: Can you provide specific activity data for my enzyme?
A: Yes, we can provide specific activity data for your enzyme. In addition to our enzyme activity assays, we also offer Sandwich ELISA assays to quantify the enzyme. By combining these two services, we can generate specific activity data, which is crucial for understanding the functional efficiency of your enzyme. -
Q: What substrates can you use in your enzyme activity assays?
A: Our assays are designed to be versatile and can accommodate a wide range of substrates, including ATP, NADH, pyruvate, and many others. By isolating the enzyme from other cellular components, we can use simple and commonly used substrates without worrying about interference from other reactions that might occur in vivo. -
Q: Can you analyze post-translational modifications (PTMs) of the enzyme during the assay?
A: Yes, we can analyze post-translational modifications (PTMs) of the enzyme. Since our immunoassay approach separates the enzyme from other cellular components, we can measure its PTMs at the time of the assay. This can be done using techniques such as mass spectrometry or by reacting the enzyme with PTM-specific antibodies, providing valuable insights into the enzyme's functional state.
-
Q: Can you customize your enzyme activity assays to meet specific research needs?
A: Yes, we can customize our enzyme activity assays to meet your specific research needs. We understand that every research project is unique, and we are committed to providing tailored solutions to help you achieve your goals. Please contact us to discuss your specific requirements, and we will work with you to design the perfect assay.
-
Q: What kind of support do you offer after the assay is completed?
A: After the assay is completed, we provide comprehensive data analysis and detailed reports to help you interpret the results. Our team is also available to answer any follow-up questions or provide additional support as needed. We aim to be a long-term partner in your research journey, ensuring that you have the resources and support you need to succeed.
Resources
Related Services
- Enzyme Immobilization
- Enzyme/Antibody conjugation
- Enzyme Target and Screening
- Drug Discovery Screening
Related Products
References:
- Guo J, Gan C, Cheng B, Cui B, Yi F. Exploration of binding mechanism of apigenin to pepsin: Spectroscopic analysis, molecular docking, enzyme activity and antioxidant assays. Spectrochimica Acta Part A: Molecular and Biomolecular Spectroscopy. 2023;290:122281. doi:10.1016/j.saa.2022.122281
- Roveri A, Di Giacinto F, Rossetto M, et al. Cardiolipin drives the catalytic activity of GPX4 on membranes: Insights from the R152H mutant. Redox Biology. 2023;64:102806. doi:10.1016/j.redox.2023.102806
Contact us or send an email at for project quotations and more detailed information.
Quick Links
-

Papers’ PMID to Obtain Coupon
Submit Now -

Refer Friends & New Lab Start-up Promotions