


Other ECM Glycoproteins
Related Symbol Search List
- THBS1
- DPT
- ECM1
- ELN
- EMILIN2
- FBLN2
- FGA
- FGB
- FGG
- Fibronectin
- LAMA4
- LAMB2
- LAMC2
- LTBP1
- LTBP2
- LTBP4
- NID1
- Nidogen-2
- POSTN
- PXDN
- SPARC
- TINAGL1
- TNC
- TNN
- Vitronectin
Immunology Background
In a multicellular organism, a complex network of cells surrounded by a variety of macromolecules, called the extracellular matrix. Studies have shown that extracellular matrices do not act as an inert support as previously thought, or connect cells together to form tissues and organs. Instead, it contains a large number of signaling molecules that actively participate in controlling cell growth, polarity, shape, migration and metabolic activity. Studies on human cells have shown that fibronectin in the extracellular matrix is mainly secreted by fibroblasts, epithelial cells, etc. and attached to the surface of the cells. Its role is to promote cell attachment to the matrix, adhesion between cells, intracellular Construction of microfilaments and stress fibers. It has been observed that the transformed in vitro cultured fibroblasts have a reduced amount of surface fibrin, and in this connection, the cell morphology is rounded, loosened with the bottom of the medium, the intracellular stress fibers are greatly reduced, and the cells are densely packed and overlapped. This transformed cell is inoculated into a normal body, often grows into a cancerous mass, invades normal tissues, and undergoes extensive metastasis.
Composition of the extracellular matrix
The composition of the extracellular matrix can be divided into three major categories: 1 glycosaminoglycans, proteoglycan, which form an aqueous gel, in which many other embeddings are embedded. Matrix components; 2 structural proteins, such as collagen and elastin, which impart a certain strength and toughness to the extracellular matrix; 3 adhesive proteins: such as fibronectin and laminin, which promote the binding of cells to the matrix. The collagen and proteoglycan are used as the basic skeleton to form a fibrous network complex on the cell surface, and the complex is directly linked to the cell surface receptor through fibronectin or laminin and other connecting molecules; or attached to the receptor on. Since the receptor is mostly a membrane integrin and is linked to the intracellular matrix protein, the extracellular matrix integrates the extracellular and intracellular cells through a membrane integrin.
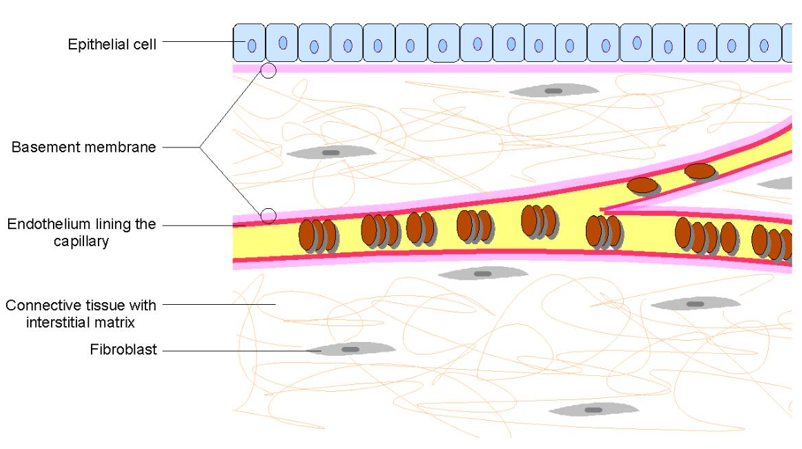 Figure 1. Architecture outside the matrix.
Figure 1. Architecture outside the matrix.
Function
Due to its diverse nature and composition, ECM can perform many functions, such as providing support, isolating tissues from each other, and regulating cell-to-cell communication. The extracellular matrix regulates the dynamic behavior of the cells. In addition, it isolates multiple cell growth factors and acts as their local storage. Changes in physiological conditions can trigger protease activity, resulting in localized release of such storage. This allows for rapid and local growth factor mediated activation of cellular functions without the need for de novo synthesis, with VEGFC (Figure 2), MMP2 and collagen I as examples. The formation of extracellular matrices is critical for processes such as growth, wound healing and fibrosis. Knowledge of the structure and composition of ECM can also help to understand the complex kinetics of tumor infiltration and metastasis in cancer biology, as metastasis usually involves enzymes such as serine proteases, threonine proteases, and matrix metalloproteinases that disrupt the extracellular matrix. The stiffness and elasticity of ECM are important in cell migration, gene expression and differentiation. The cells actively sense the rigidity of the ECM and preferentially migrate to the hard surface. This phenomenon is called durotaxis. They also detect elasticity and regulate their gene expression accordingly, and they have increasingly become the subject of research due to their effects on differentiation and cancer progression.
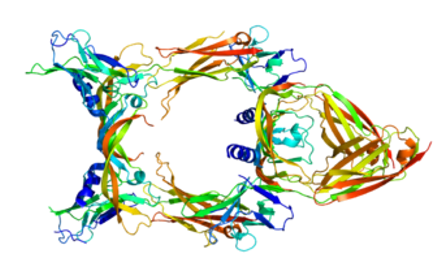 Figure 2. Structure of protein VEGFC
Figure 2. Structure of protein VEGFC
Physiology
1. Effect on gene expression
Different mechanical properties in ECM affect both cell behavior and gene expression. Although the mechanism of its completion has not been fully explained, it is believed that the adhesion complex and the cytoskeleton of actin-myosin, whose contractile force is transmitted through the transcellular structure, play a key role in the molecular pathways that have not yet been discovered.
2. Effect on differentiation
ECM elasticity can direct cell differentiation, a process in which cells change from one cell type to another. In particular, naive mesenchymal stem cells (MSCs) have shown specific lineages and have a phenotype that is extremely sensitive to tissue-level elasticity. MSCs are placed on soft matrices that mimic the brain's differentiation into neuron-like cells, which have similar shapes, RNAi profiles, cytoskeletal markers, and transcription factor levels. Similarly, the harder matrix that mimics muscle is myogenic, while the rigid matrix that mimics collagenous bone is osteogenesis.
References:
1. Jeltsch M.; et al. Hyperplasia of lymphatic vessels in VEGF-C transgenic mice. Science. 1997, 276 (5317): 1423–1425.
2. Joukov V.; et al. A novel vascular endothelial growth factor, VEGF-C, is a ligand for the Flt4 (VEGFR-3) and KDR (VEGFR-2) receptor tyrosine kinases. The EMBO Journal. 1996,15 (2): 290–298.